Giovanni Chetta 박사 편집
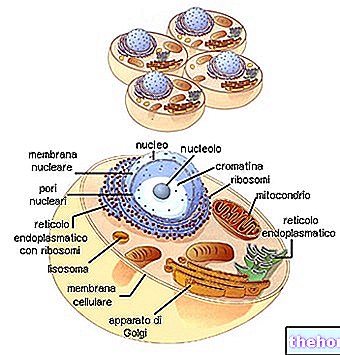
ECM은 일반적으로 몇 가지 큰 종류의 생체 분자로 구성되는 것으로 설명됩니다.
- 구조 단백질(콜라겐 및 엘라스틴)
- 특수 단백질(피브릴린, 피브로넥틴, 라미닌 등)
- 프로테오글리칸(아그레칸, 신데칸) 및 글루사미노글리칸(히알루로난, 콘드로이틴 설페이트, 헤파란 설페이트 등)
구조 단백질
콜라겐은 동물계에서 가장 대표적인 당단백질 계열을 형성합니다. 그들은 세포외 기질에 가장 많이 존재하는 단백질(가장 중요한 것은 아님)이며 적절한 결합 조직(연골, 뼈, 근막, 힘줄, 인대)의 기본 구성요소입니다.
콜라겐에는 최소 16가지 유형이 있으며, 그 중 유형 I, II 및 III는 전형적인 원섬유 수준에서 가장 많이 존재합니다(유형 IV는 기저판의 주요 구성요소를 나타내는 일종의 세망을 형성함).
콜라겐은 대부분 섬유아세포에 의해 합성되지만 상피세포도 합성할 수 있습니다.
콜라겐 섬유는 세포의 생명에 필수적인 생물학적 연속체를 구성하는 엄청난 양의 세포외 기질의 다른 분자와 지속적으로 상호 작용합니다.피브릴의 관련 콜라겐은 장력에 저항할 수 있는 구조의 형성 및 유지에 주된 역할을 합니다. 거의 비탄력적입니다(글루코사밍글리칸은 압축에 대한 저항 작용을 수행합니다.) "점탄성 근막"은 사람의 자세에 큰 영향을 미칩니다. 예를 들어, 환경 영향에 따라 콜라겐이 변화하는 능력의 추가 증명으로 다양한 정도의 강성, 탄력성 및 저항을 감안할 때 프로테오글리칸("글루코사미노글리칸 및 프로테오글리칸" 단락에 설명됨)처럼 기능적으로 기능할 수 있는 FACIT(Fibril Associated Collagen with Interrupted Triple helix)라는 용어로 정의된 콜라겐이 있습니다.
콜라겐 섬유는 PG/GAG(프로테오글리칸/글루코사미노글리칸) 코팅 덕분에 바이오센서와 생체전도체의 특성을 가지고 있습니다.
구조적 변형을 생성할 수 있는 모든 기계적 힘은 분자간 결합에 응력을 가하여 약간의 전기 플럭스를 생성한다는 것을 알고 있습니다. 압전 전류 (Athenstaedt, 1969). 이러한 경우 콜라겐 섬유는 볼록면에 양전하를, 오목면에 음전하를 분포시켜 반도체로 변형됩니다(일방향 표면에서 전자의 흐름을 허용함). 압전 에너지(열 스트레스에 의해 생성된 초전 에너지)는 매우 짧은 시간(약 10-7~10-9초)에 순환하는 이온에 의해 중화되기 때문에 신호 상의 PG/GAG 배열 전기 임펄스의 "중계기"로 작용하는 것과 같이 섬유소 표면의 신호 전파에 결정적입니다. 특히, 대략적인 종주기. 64 nm(광학 현미경 아래에서 줄무늬로 나타남)는 약 64 m/s(고속 신경 섬유의 전도 속도에 해당)에 해당하는 임펄스의 전파 속도를 허용합니다. - Rengling, 2001. 콜라겐 원섬유의 강한 쌍극자 모멘트 MEC의 낮은 유전상수뿐만 아니라 공명 용량(모든 펩타이드 구조에 공통적인 특성)은 전자기 신호의 전송을 용이하게 합니다.따라서 3차원 및 유비쿼터스 콜라겐 네트워크는 또한 생체 전기 신호를 전도하는 독특한 특성을 가지고 있습니다. 구심성 방향(ECM에서 세포로) 또는 그 반대로 원심성 방향으로 콜라겐 원섬유와 세포 사이의 상대적인 배열을 기반으로 한 공간의 3차원.
이 모든 것은 실시간 MEC-세포 통신 시스템을 나타내며 이러한 전자기 생체 신호는 중요한 생화학적 변화를 유발할 수 있습니다. 예를 들어 뼈에서 파골세포는 압전하를 띤 뼈를 "소화"할 수 없습니다(Oschman, 2000).
마지막으로, 세포는 놀랍지 않게 지속적으로 상당한 에너지 소비(약 70%)와 함께 반드시 배출되어야 하는 물질을 생산한다는 점을 강조해야 합니다. 이 물질은 주로 특정 세포에서 프로토콜라겐(콜라겐의 생물학적 전구체)의 독점적인 저장을 통해 배출되어야 합니다. 소포(Albergati, 2004).
대부분의 척추동물 조직은 힘과 탄력이라는 두 가지 중요한 특성이 동시에 존재해야 합니다. 이러한 조직의 ECM 내부에 있는 실제 탄성 섬유 네트워크는 강한 견인 후 초기 상태로 돌아갈 수 있도록 하며, 탄성 섬유는 장기 또는 그 일부의 확장성을 5배 이상 증가시킬 수 있습니다. 긴 비탄성 콜라겐 섬유가 탄성 섬유 사이에 산재되어 "조직의 견인으로 인한 과도한 변형을 제한하는 정밀한 작업. L"엘라스틴 탄성 섬유의 주성분을 나타냅니다. 콜라겐은 프롤린과 글리신이 풍부하지만 콜라겐과 달리 당화되지 않고 하이드록실리신이 아닌 하이드록시프롤린 잔기를 많이 포함하기 때문에 아미노산 길이가 약 750개 정도의 극소수성 단백질입니다. 엘라스틴은 모든 결합 조직의 ECM을 관통하는 섬유와 라멜라로 구성된 불규칙한 3차원 형태의 실제 생화학적 네트워크로 나타나며, 특히 탄성 특성을 가진 혈관에서 풍부한 양으로 발견됩니다(더 많은 ECM의 단백질입니다. 동맥, 인대, 폐 및 피부에 존재하며 대동맥의 총 건조 중량의 50% 이상을 차지합니다. 진피에서는 콜라겐과 달리 엘라스틴의 밀도와 부피가 시간이 지남에 따라 증가하는 경향이 있지만 오래된 엘라스틴은 일반적으로 부풀어 오르고 거의 부풀어 오르며 종종 조각난 모양과 구성 요소의 감소로 나타납니다. (Pasquali Rochetti et al, 2004). 평활근 세포와 섬유아세포는 세포외 공간에서 분비되는 전구체인 트로프엘라스틴의 주요 생산자입니다.
"콜라겐과 엘라스틴, 세포외 기질의 콜라겐 섬유"에 대한 다른 기사
- 세포외 기질
- 피브로넥틴, 글루코사미노글리칸 및 프로테오글리칸
- 세포 평형에서 세포외 기질의 중요성
- 세포외 기질 및 병리의 변경
- 결합 조직 및 세포외 기질
- 깊은 근막 - 결합 조직
- 근막 기계수용체 및 근섬유아세포
- 깊은 근막 생체 역학
- 자세와 동적 균형
- 텐세그리티와 나선운동
- 하지와 몸의 움직임
- 둔근 지지대 및 구강악 장치
- 임상 사례, 자세 변화
- 임상 사례, 자세
- 자세 평가 - 임상 사례
- 참고 문헌 - 세포외 기질에서 자세까지. 연결 시스템이 우리의 진정한 Deus ex machina입니까?