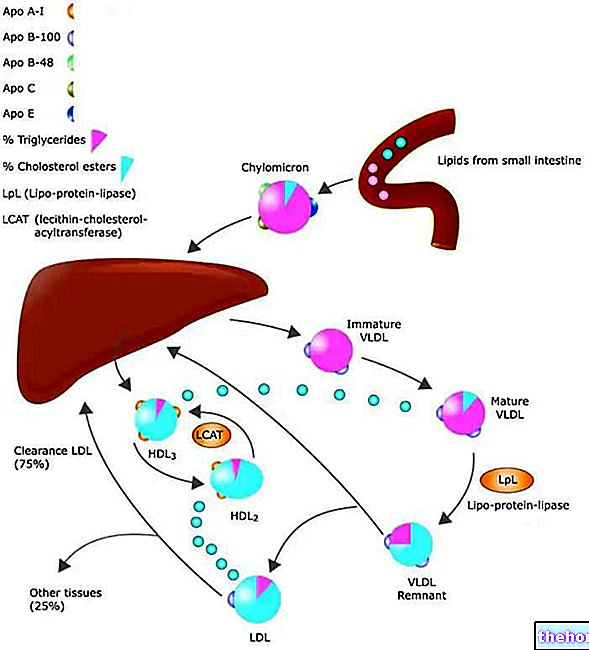
크렙스 회로는 트리카르복실산 회로라고도 하며 해당 과정에 의해 생성된 피루브산에 대한 피루브산 탈수소효소의 작용에 의해 얻어지는 아세틸 코에짐 A를 출발 대사물로 사용합니다.
ATP와 환원력은 크렙스 사이클에서 얻습니다. 환원력은 NADH와 FADH2가 각각 NAD+와 FAD로 산화되는 호흡 사슬로 보내집니다. 환원력은 호흡 사슬을 따라 커플링 시스템으로 전달되어 더 많은 ATP가 생성됩니다.
크렙스 회로는 포도당 대사뿐만 아니라 지방산과 아미노산의 대사를 위한 신경 중추입니다. 사실 아세틸 조효소 A로 전환되는 피루브산은 포도당 분해에서만 오는 것이 아닙니다. 예를 들어, 또한 알라닌(아미노산)의 아미노기 전이로부터.
크렙스 회로에 참여하는 아세틸 조효소 A의 약 80%는 지방산 대사에서 나옵니다.
Acetyl coenzyme A는 thioester이므로 높은 에너지 함량을 가지고 있습니다. 구연산 합성 효소 새로운 탄소-탄소 결합을 형성하기 위해; 구연산 합성 효소는 크렙스 회로의 첫 번째 효소입니다.
아세틸 조효소 A의 메틸 탄소는 (호변이성화에 의해) 양성자를 자발적으로 방출하고(탄소음이온이 됨) 옥살로아세테이트의 카르보닐 탄소를 공격합니다. 고에너지 함량의 티오에스테르(시트릴 조효소 A)가 형성되며, 이로부터 가수분해에 의해 시트레이트가 얻어집니다. 그리고 코엔자임 A는 재형성됩니다. 구연산염 합성효소는 생성물, 즉 구연산염과 ATP에 의해 부정적으로 조절됩니다. 변조기).

ATP는 또한 ATP가 생성되는 호흡 사슬로 보내지는 크렙스 회로에서 환원력이 얻어지기 때문에 구연산 합성 효소의 작용에 영향을 미칩니다. ATP가 축적되면 필요한 것보다 더 많이 생산된다는 의미입니다. 크렙스 주기를 늦춤으로써(단계 중 하나가 느려지면 주기가 느려짐) ATP 생성도 느려집니다. ATP의 음의 조절은 피드백 조절입니다(최종 생성물 중 하나의 형성은 프로세스에서 한 단계의 속도를 조정하여 변조됨).
크렙스 회로의 두 번째 단계에서 구연산염은 효소의 작용에 의해 이소시트르산으로 전환됩니다. 아코니타제; 효소의 이름은 시트레이트가 먼저 시스-아코니테이트의 형성으로 탈수되고, 이어서 물이 이전에 결합된 탄소와 다른 탄소에 부착되어 다시 들어간다는 사실에서 유래합니다. 이소시트레이트는 기질이 촉매 부위를 떠나지 않고 얻어지며, 아코니타제는 입체특이성 효소입니다: 이것은 시트레이트의 3개의 카르복실 중심을 인식하고 이것은 시트레이트가 효소에 결합된 상태로 남아 있게 하여 "물"의 출구와 입구가 항상 통과하도록 합니다. cis-aconitate 중간체를 통해.
크렙스 주기의 세 번째 단계에서는 이산화탄소로 제거되는 탄소의 손실이 있기 때문에 첫 번째 에너지 고려 사항이 있습니다. 이 단계를 촉매하는 효소는 이소시트레이트 탈수소효소; 기질은 우선 탈수소화를 거칩니다: NAD +는 환원력을 획득하고 옥살로숙시네이트가 형성됩니다(이것은 숙신산의 옥살 유도체임).옥살로숙시네이트는 그 다음 α-케토글루타레이트로 탈카르복실화됩니다.
효소 isocitrate dehydrogenase에는 두 개의 조절 부위가 있습니다: ADP로 인한 양성 조절과 ATP로 인한 음성 조절. 매일 소비되는 ATP의 양은 매우 높습니다: ATP는 가수분해, "ADP 및 모든" 오르토포스페이트에 의해 방출되는 에너지를 제공합니다.
유기체에서 뉴클레오사이드(질소 염기와 당) 및 뉴클레오타이드(뉴클로사이드와 인산염)의 총 농도는 거의 일정합니다. ATP는 거의 없음) ADP는 에너지 필요의 동의어이므로 긍정적인 조절자이고 ATP는 에너지 가용성의 증상이므로 부정적인 조절자입니다.
계속: 두 번째 부분 "

.jpg)