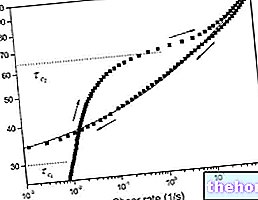
세포가 혐기성 조건에서 작동하면 포도당을 젖산으로 전환하여 에너지를 생성하고 Cori 회로를 통해 젖산을 폐기합니다. 나머지 10%만 혐기성으로. 유산소 경로가 제공할 수 있는 것보다 더 많은 ATP가 필요한 경우(예: 근육이 스트레스를 받을 때) 혐기성 대사에 의해 추가 공급이 제공됩니다(우리는 산소 결핍 상태에 있습니다: 숨가쁨, 피로 호흡 등): 포도당 신생합성을 통해 젖산(해당 분해에서 얻음)을 포도당으로 전환하여 이 대사를 가속화하는 것이 필요합니다.
호기성 대사는 미토콘드리아에서 발생합니다.
호기성 대사에서 가장 먼저 만나는 효소는 피루브산 탈수소효소; 피루브산 탈수소효소는 3개의 촉매 부위가 연속적으로 작용하는 48~60개의 단백질 단위의 집합체이기 때문에 효소라기 보다는 효소 복합체라고 말하는 것이 더 정확합니다.
피루브산 탈수소효소는 다음 반응(산화환원)을 촉매합니다.
피루브산 + NAD + + CoA-SH → 아세틸 CoA + NADH + H + + CO2
CoA-SH는 조효소 A: 판토텐산의 유도체, 아세틸 조효소 A는 티오에스테르입니다. 이것은 피루브산의 첫 번째 탄소가 산화수 3에서 산화수 4로 가고(산화됨) 피루브산의 두 번째 탄소가 산화수 2에서 산화수 3(산화됨)으로 가기 때문에 산화환원 과정입니다. 그런 다음 피루브산은 산화되고(두 개의 전자를 모두 잃음) NAD는 환원됩니다.
언급한 바와 같이, 피루브산 탈수소효소는 3가지 유형의 효소 활성을 가지며, 각각은 자체 촉매 보조인자에 의해 지원됩니다.
- 티아민 피로포스페이트(비타민 B1의 유도체); 그것은 탈양성자 형태로 활성화되어 있습니다. 탄소 음이온이 형성됩니다.
- 리포아미드(리포산의 유도체), 반응성이 매우 높은 이황화 다리를 포함합니다.
- 플라빈 아데닌 디뉴클레오티드(비타민 B2의 유도체); 그것은 산화 환원 특성을 가진 뉴클레오티드입니다. 산화 환원 중심은 플라빈으로 구성됩니다.
진핵 세포에서 호기성 대사는 미토콘드리아인 세포의 특수 소기관에서 발생합니다. 박테리아에서 포도당과 다른 종의 대사는 세포에서 일어나지만 전문화된 세포 소기관은 없습니다.
피루브산이 미토콘드리아에 들어갈 때, 포도당신생합성(시작 물질을 재구성하기 위해)을 수행할 필요가 있는 경우 "피루브산 카르복실라제의 작용을 받거나, 에너지 생성이 필요한 경우 피루브산 탈수소효소의 적용을 받을 수 있습니다. " 호기성 대사에 의해 생성되는 아세틸 조효소 A는 pyruvate carboxylase의 작용을 자극하여 글루코스 신생합성을 촉진하고 pyruvate dehydrogenase의 작용을 감소시킨다.
이제 피루브산 탈수소효소가 어떻게 작용하는지 봅시다. 우선, 티아민 피로포스페이트의 작용에 의한 피루브산의 탈카르복실화가 있습니다.
산성 환경은 티아민 피로포스페이트의 음이온 형태가 활성이고 산성 pH에서 양성자화되고 탈카르복실화가 일어나지 않기 때문에 호기성 대사를 억제할 수 있습니다.
탈카르복실화는 탄소-탄소 결합이 끊어져야 하기 때문에 어려운 반응입니다. 이 경우 반응 중간체(하이드록시에틸-티아민 피로포스페이트)가 공명(분자의 p-전자가 비편재화됨)을 제공한다는 사실에 의해 반응이 열역학적으로 유리합니다. 하이드록시에틸-티아민 피로포스페이트는 세 가지 가능한 형태(공명)로 존재하고 이것은 그것을 상당히 안정적으로 만듭니다. 또한 음이온 형태의 hydroxyethyl-thiamine pyrophosphate는 lipoamide의 disulfide bridge(pyruvate dehydrogenase의 두 번째 촉매 보조인자)와 상호작용할 수 있을 만큼 충분히 오래 생존합니다. 길고 유연한 사슬의 끝) 효소 복합체의 한 촉매 부위에서 다른 촉매 부위로 이동할 수 있습니다.
그런 다음 리포아미드는 이황화 다리를 통해 히드록시에틸-티아민 피로포스페이트에 결합합니다: 아세틸 리포아미드가 얻어집니다. 이것은 피루브산 탈수소효소 복합체의 첫 번째 효소에 의해 촉매되는 트랜스아세틸화 반응의 첫 번째 단계입니다. 원래의 형태로 되돌아간 히드록실기와 티아민 피로포스페이트: 이황화 다리가 아세틸로 산화된 히드록실기에 대해 산화제(두 개의 황 원자가 환원됨)로 작용하는 산화환원 반응이 발생했습니다.
이 단계 후에, 리포아미드의 진동 팔은 아세틸 그룹을 운반함으로써 진정한 트랜스아세틸라제 활성을 수행하는 피루브산 탈수소효소의 두 번째 효소로 이동하여 접근합니다. 두 번째 효소에 의해 촉매되는 트랜스아세틸화 반응의 두 번째 단계가 발생합니다. 이러한 방식으로 우리는 아세틸 조효소 A를 얻었습니다. 이제 환원된 형태의 리포아미드를 복원할 필요가 있습니다: 피루브산 탈수소효소의 세 번째 효소가 개입하여 리포아미드를 산화환원하고 전극을 FADH2로 환원되는 FAD로 전달합니다. FAD/FADH2는 두 개의 별개의 단일전자 단계 또는 단일 생체전자 단계에서 산화환원 쌍으로 기능할 수 있습니다.
FADH2는 즉시 전자를 NAD +에 제공하여 FAD 및 NADH + H +를 얻습니다.
기술된 바와 같이 얻은 아세틸 조효소 A는 크렙스 회로(또는 트리카르복실산의 회로)의 출발 산물입니다.













-cloruro.jpg)