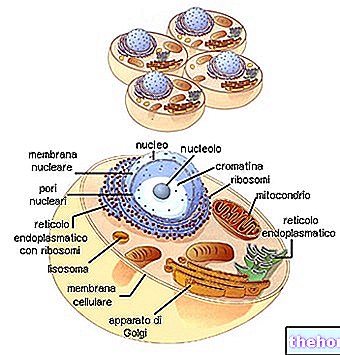
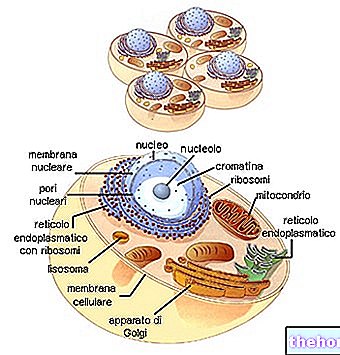
유사 분열은 일반적으로 각각 prophase, metaphase, anaphase 및 telophase라고하는 4 개의 기간으로 나뉩니다. 그런 다음 세포분열이라고 하는 두 개의 딸세포로 분열됩니다.
제안
핵에서는 여전히 가늘고 긴 공 모양으로 감긴 채색 가능한 필라멘트의 윤곽을 점차적으로 볼 수 있습니다. 따라서 핵 단백질에 결합된 DNA 가닥의 점진적인 나선화는 염색체를 점차적으로 식별 가능하게 만듭니다. 한편 핵소체는 사라지고 중심소체는 두 배가 됩니다. 두 중심소자는 핵막의 용해가 시작되는 동안 핵의 반대 극으로 이동합니다. prophase에서 metaphase로 넘어가는 순간(일부는 prometaphase로 별도로 식별하기 때문에) 염색체가 짧아지고 명확하게 보이며 더 이상 핵막에서 분리되지 않습니다. 중심 소체는 반대 극에 있으며 자오선 방식으로 연결하는 미세 소관의 방추와 함께 핵소체가 용해되었습니다. 무채색 방추라고 하는 중심소를 연결하는 방추는 염색할 수 없기 때문에(염색체와 달리) 연속 섬유(방추 섬유)와 중간점에서 염색체의 중심에 연결되는 섬유(염색체 섬유)를 모두 포함합니다.

중기
중기에서 방추는 명확하게 구별되며 모든 염색체가 적도 판이라고 불리는 적도 평면에 배열됩니다. 이 시점에서 염색체는 최대 단축 상태에 있습니다. 이것은 세포가 염색체를 세고 식별하기 위해 고정되는 순간입니다. 각 염색체는 명확하게 두 개의 동일한 필라멘트(염색분체)로 구성되어 있으며 중심체( 두 염색분체는 복제의 결과입니다.) 중심체는 두 혈소판 사이의 일종의 접착과 같은 유일한 접촉점입니다. 모든 중심체는 무채색 방추의 염색체 섬유의 중심점에 부착되어 있습니다(이것이 염색체가 적도 위치에 있는 이유입니다).
아나페이즈
중기의 끝에서 각 중심체는 두 배가 되며 각 반쪽은 각 극 방향으로 방추를 따라 이동합니다. 이 시점에서 각각의 중심체에 의해 끌린 염색분체는 두 그룹으로 명확하게 분리됩니다. 이렇게 분리된 각 염색분체는 말하자면 성숙합니다. 이제부터는 각각의 딸 세포로 향하는 염색체입니다.
말기
2개의 동일한 클러스터로 분리된 염색체는 2개의 새로운 세포의 핵을 재구성하여 탈감기로 되돌아갑니다. 무채색 용융물이 용해됩니다.
세포분열
세포질은 또한 두 개의 딸 세포가 부피와 세포 소기관의 각각의 비율에 기인하는 점진적 조절에 의해 분할됩니다. 특히, 각 딸세포는 중심소체(이들은 언급한 바와 같이 고유한 유전적 연속성을 가진 구조임)를 받았기 때문에 최소한 하나의 미토콘드리아를 받아야 합니다.
식물성 왕국에서는 유사 분열의 일반적인 특징이 존중되지만 몇 가지 차이점이 있음에 유의해야합니다. 우선, 중심소가 없습니다. 방추의 극에는 중심체라고 불리는 광학적으로 빈 공간이 있으며, 이 공간에서 미세 소관이 방출됩니다. 또한, 각 딸세포에 대해 색소체의 귀인(유전적 연속성으로 인해)도 보장되어야 하는 세포분열시 딸세포의 분리는 교살에 의한 것이 아니라 격막의 형성에 의해 발생하며, 처음에는 원형질막만 발생하고 그 다음에는 세포벽이 삽입됩니다.
유전적 기초는 "유전 물질의 배가(DNA의 복제, 즉 각 염색체가 중심체를 통해 연결된 두 개의 동일한 염색분체로 두 배로 증가)와 반감기(중심체의 분리, 이동 2개의 새로운 동일한 핵을 구성하기 위해 반대 방향으로 2개의 염색분체).
우리가 볼 수 있듯이 염색체는 상동체 쌍으로 존재하기 때문에 (각각 배우자에서 유래) 분열 직후 염색체 가닥의 수가 각 유형의 염색체에 대해 한 쌍임을 알 수 있습니다. 단일 종의 다른 유형의 염색체 특성의 수를 n이라고 하면 유사분열 후 정상적인 염색체 세트는 n쌍의 상동 염색체(2n 염색체 = 이배체 세포)입니다.
그러나 S기 이후에는 각 염색체가 두 배가 됩니다. 사실, 각 딸세포에 2n개의 염색체를 줄 수 있으려면 4n개의 염색분체가 있어야 합니다. 이런 식으로 우리는 복제와 유사 분열이 DNA의 4n 가닥과 2n 가닥 사이에서 교대로 일어나는 것을 볼 수 있습니다.